
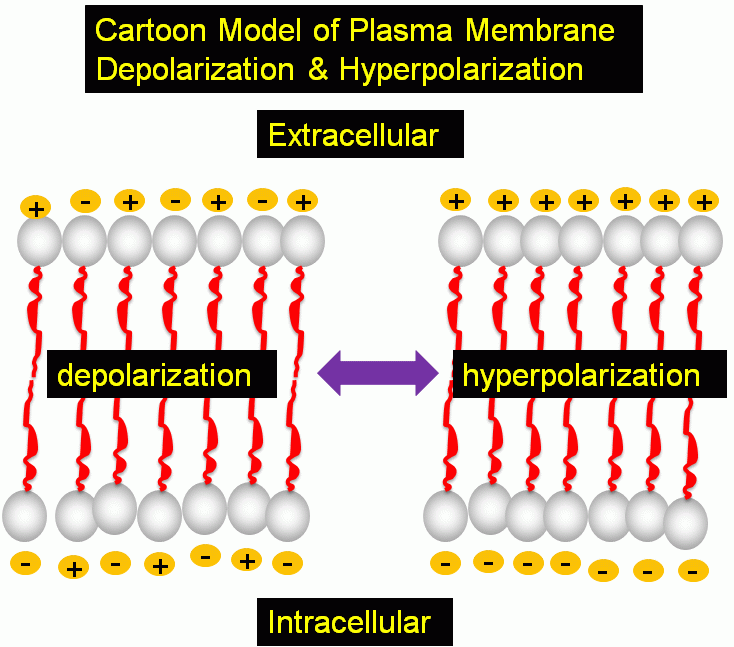
The plasma membrane of mammalian cells has a transmembrane potential of -70mV (negative inside) due to the distribution of K+, Na+ and Cl- ion gradients which are maintained by the process of active transport across the plasma membrane. Changes in potential difference across the plasma membrane plays a role in many phsiological processes. Plasma membrane hyperpolarization and depolarisation, or increase and decrease in membrane potential due to fluctuations in ion concentrations can be monitored by a range of fluorescent dyes.
Cationic Carbocyanine dyes such as the DiO series label all cell membranes but at low concentrations (4-40nm) label only mitochondria. However, anionic Oxonol dyes label only the plasma membrane of cells making it possible to measure hyperpolarization and depolarisation of the plasma membrane during physiological processes kinectically by flow cytometry.
The dye bis-(1,3-dibutylbarbituric acid)pentamethine) oxonol or DiBAC4(5) is excited at 590nm and emits at 616nm and thus can be used in flow cytometers fitted with an argon laser (488nm) with the signal collected in a standard PE filter setup (575/26nm) or PE-Texas Red (610/10nm).
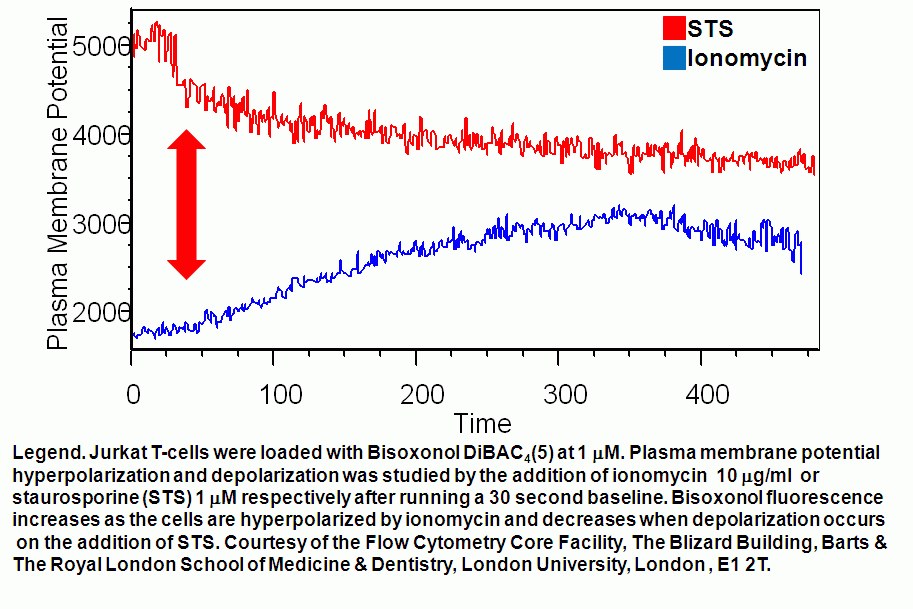
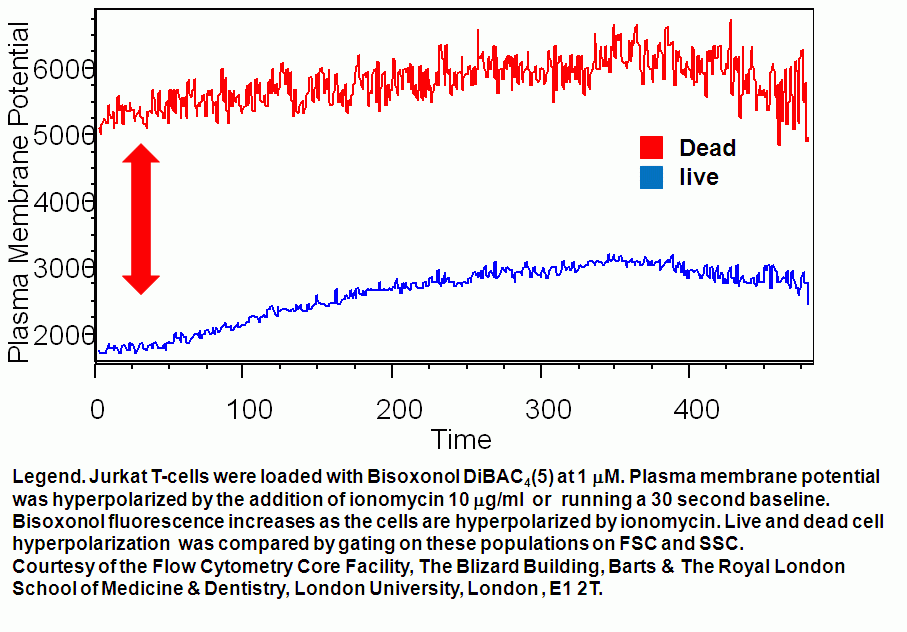
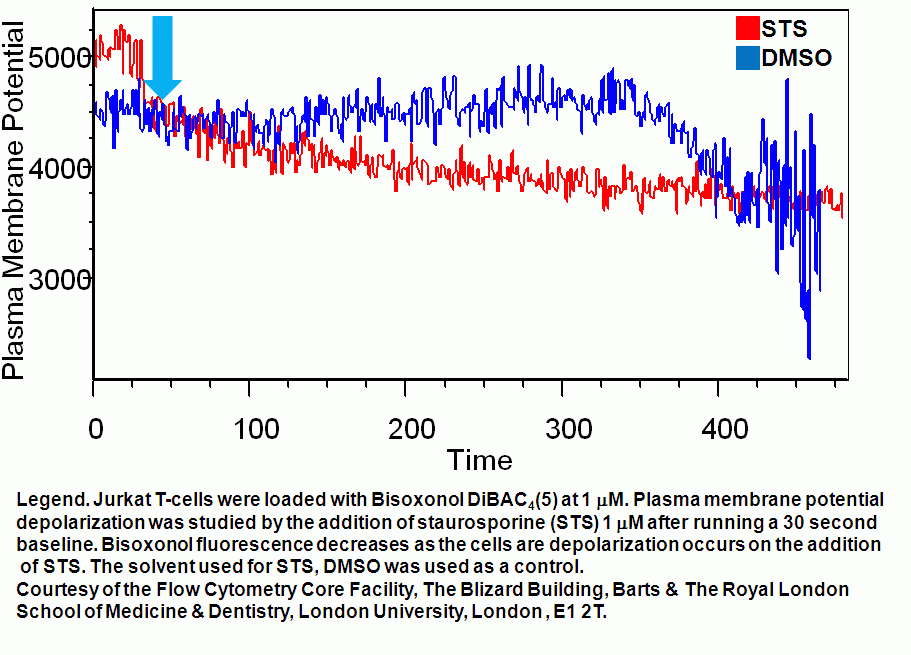
Ionomycin can be used to depolarize the plasma membrane of live T cells. DMSO and ethanol (in small volumes) can be used as controls for such experiments. While Staurosporine (STS) causes live T cell plasma membranes to hyperpolarize.
- Model of plasma membrane transmembrane potential
- Plasma membrane potential hyperpolarization & depolarization kinetics
- Plasma membrane potential depolarization differences in live and dead T cells
- T cell plasma membrane hyperpolarization kinetics
- Protocol

{kind=link}