
Organelle function - mitochondrial membrane potential (Δψm) changes
The Δψm is due to a differential distribution of proteins on either side of the impermeable inner mitochondrial membrane. In apoptosis, irrespective of the stimulus, loss of Δψm occurs with the formation of a mitochondrial permeability transition pore (PT) or mitochondrial megachannel.
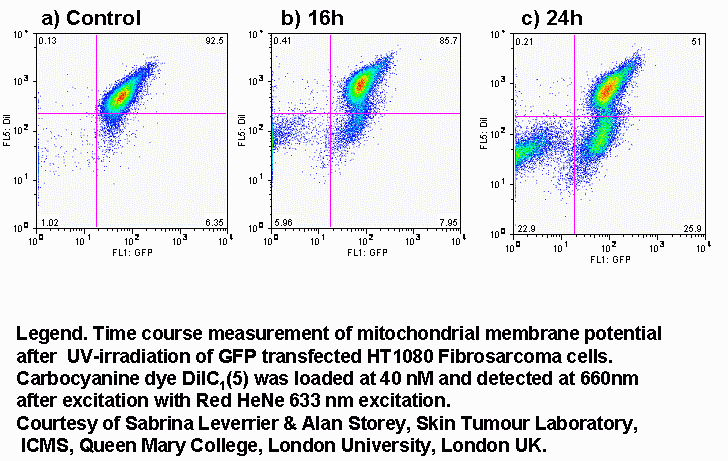
The fall in mitochondrial inner membrane potential can be measured flow cytometrically. Normal mitochondrial function has a requirement for a negative charge on the inner side of the mitochondrial inner membrane. This is maintained by the asymmetrical distribution of H+ ions across the membrane giving rise to both a potential difference and a chemical pH gradient. Cationic lipophilic dyes such as CMXRos (chloromethyl-X-rosamine), TMRE (tetramethylrhodamine), JC-1, DiOC6(3), DiIC1(5) and rhodamine 123 localize across the inner mitochondrial membrane a process which is dependent upon H+ distribution.
Loss of mitochondrial membrane potential is detected in cells by measuring a loss of dye from the inner mitochondrial matrix or a change in fluorescence of the dye as the transmembrane potential drops.
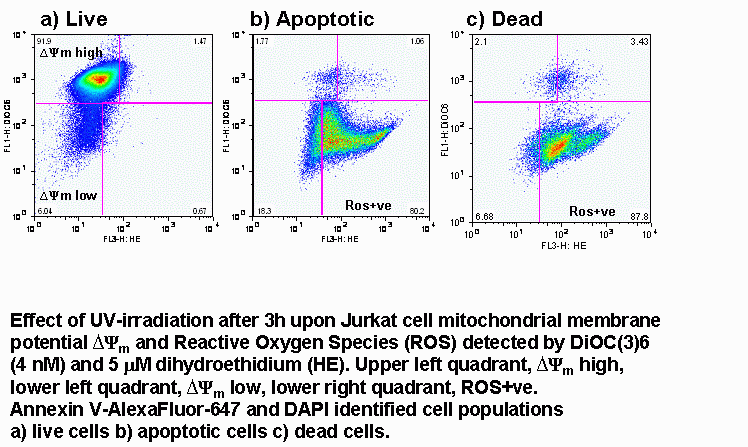
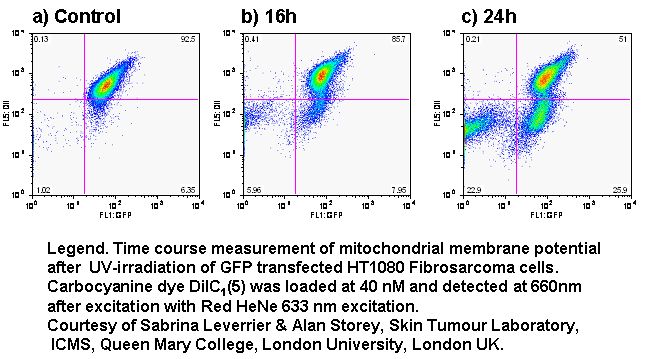
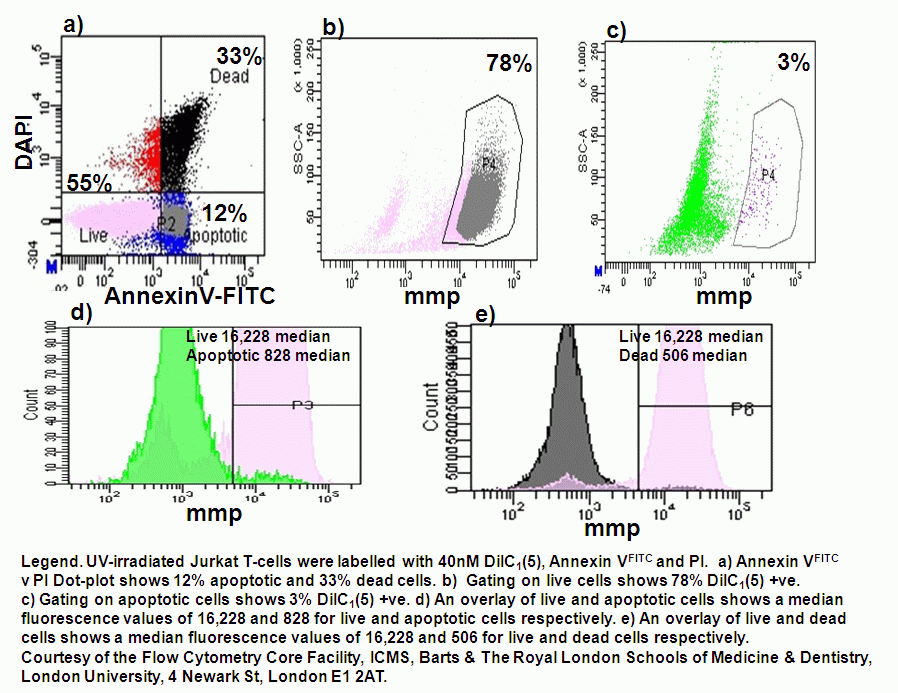
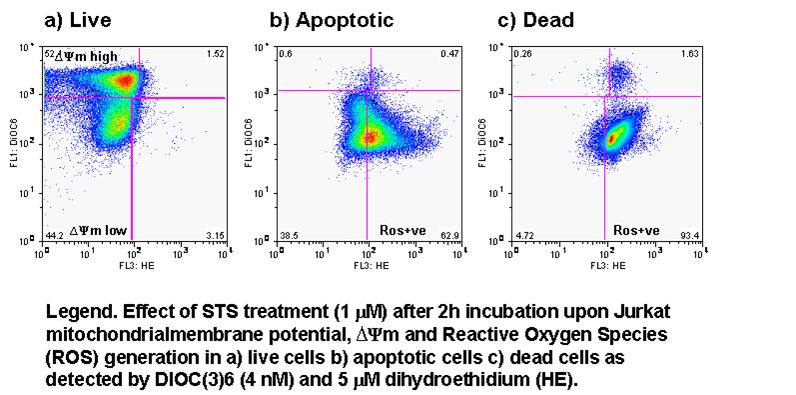
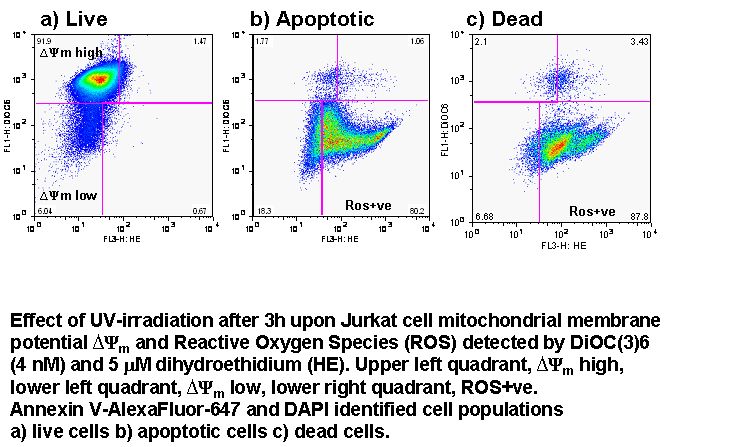
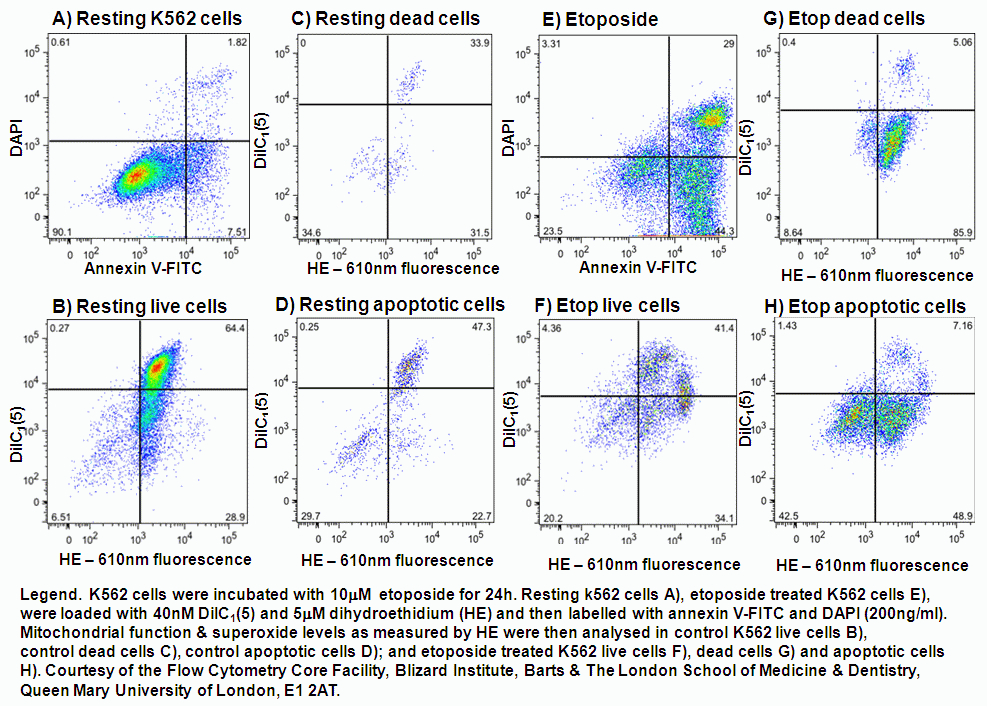
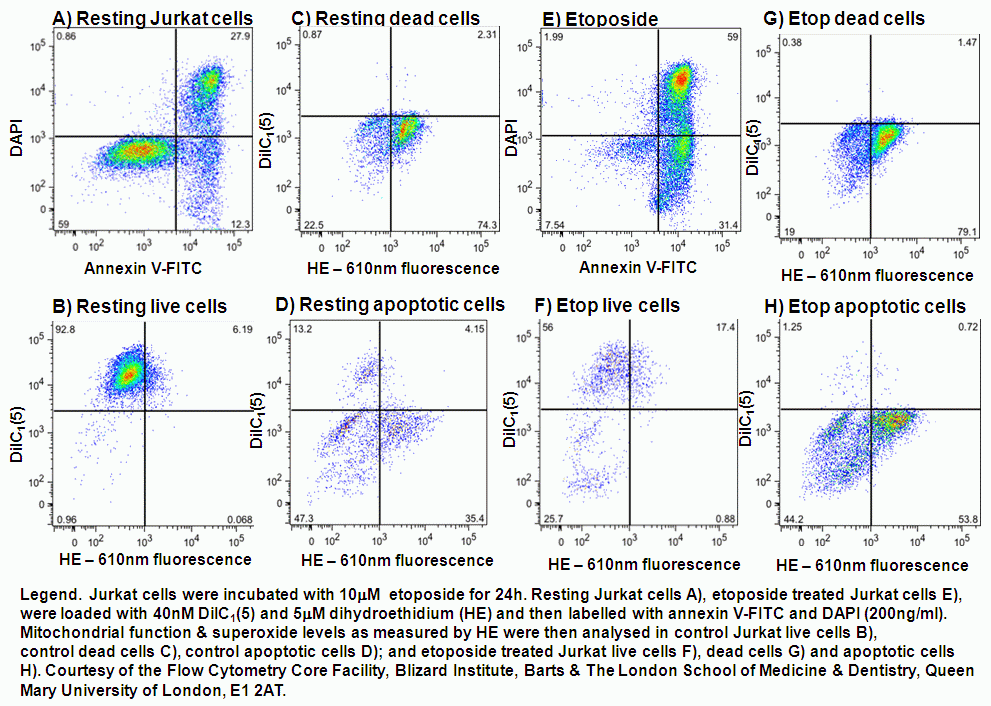
UV-irradiation, staurosporine and etoposide induction of apoptosis causes Jurkat cells to lose mitochondrial function over time. Labelling such cells with annexin V-FITC, DAPI and DiIC(5) shows that not only do apoptotic cells lose mitochondrial function a high proportion of live cells progressively lose mitochondrial function with increasing exposure to staurosporine, see figure.
e.g. DiOC6(3), DiIC1(5) with GFP expression and viability
{kind=link}
{kind=link}
{kind=link}
{kind=link}